| Vol.45, No.4, Octubre-Diciembre, 75-84, 2018 CE: 1737 CF: cag104182202 Revista Centro Agrícola Universidad Central “Marta Abreu” de las Villas ISSN papel: 0253-5785 ISSN on line: 2072-2001 |
|---|
ARTÍCULO DE INVESTIGACIÓN
Evolución de lateralización poblacional en Dysdercus andreae L. (Heteroptera: Pyrrhocoridae)
Evolution of population level lateralization in Dysdercus andreae L. (Heteroptera: Pyrrhocoridae)
Alán Rivero-Aragón1*, Ana María Suárez-Hernández1, Armando Alvaré-Jaramillo1
1 Departamento de Biología, Facultad de Ciencias Agropecuarias, Universidad Central "Marta Abreu" de las Villas (UCLV). Carretera a Camajuaní km 5 ½. Santa Clara, Villa Clara, Cuba, CP 54830
*Autor para correspondencia: alanra@uclv.edu.cu
RESUMEN
Los insectos son excelentes modelos para probar las hipótesis que proponen que la evolución de la lateralización y la simetría direccional a escala de población, son características solo de especies sociales cooperativas. El objetivo principal de este trabajo fue comprobar la existencia de lateralización poblacional en Dysdercus andreae L. (Heteroptera: Pyrrhocoridae). Desde el punto de vista teórico es interesante, considerando que esta especie forma grupos de varios cientos de individuos, lo que indica cierta coordinación, pero no es estrictamente una especie social. Para dar respuesta al objetivo trazado se construyeron dos laberintos conductuales en "T", uno para evaluar la respuesta con integración neural y otro para evaluar reflejo de escape. Se determinó que existe una conducta poblacional lateralizada en Dysdercus andreae. Se comprobó que la respuesta de escape y las decisiones sin estrés son comportamientos que evolucionan separadamente en la especie: los individuos prefieren la izquierda al reaccionar con integración neural, mientras que al responder de modo reflejo a estímulos aversivos escapan indistintamente a la derecha o izquierda. Los resultados muestran también que existe relación entre la dirección de respuesta sin estrés y la dirección de escape. Lo que puede ser indicativo de como la lateralización poblacional, existente en un inicio para respuestas más complejas, influye progresivamente sobre la conducta intuitiva de escape, conduciendo también hacia la lateralización poblacional de esta.
Palabras clave: asimetría, comportamiento animal, insectos, invertebrados, lateralidad, zurdo
ABSTRACT
The hypothesis that propose the evolution of lateralization and directional symmetry as features of cooperative social species, can be tested with insects models. The main objective of this study was to verify the existence of population level lateralization in Dysdercus andreae L. (Heteroptera: Pyrrhocoridae). This species forms groups of several hundred individuals, indicating some coordination, but it is not strictly a social species. We constructed two behavioral T-mazes to respond to the stated objective, one to evaluate response with neural integration and another to evaluate escape reflex. The results show that there are lateralized population behavior and directional symmetry in Dysdercus andreae. In addition, the escape response and decisions without stress are behaviors that evolve separately in the species; individuals prefer the left to react with neural integration. In the other hand, the insects react to aversive stimuli escaping indistinctly to the right or left. Besides, there is a relationship between the direction of response without stress and the direction of escape. It can be seen, as an indicative that the population level lateralization, developing at the beginning for more complex responses, progressively influences the intuitive escape behavior, leading to the population level lateralization of this behavior.
Keywords: asymmetry, animal behavior, insect, invertebrates, laterality, handedness
INTRODUCCIÓN
El neurólogo francés Pierre Paul Broca (Broca, 1865) describió uno de los descubrimientos más relevantes acerca de las funciones cerebrales cuando dijo: "Nous parlons avec l'hémisphère gauche!" ("Hablamos con el hemisferio izquierdo"). A partir de este trabajo se sucedieron un sinnúmero de investigaciones en el campo de la lateralización de las funciones en el Sistema Nervioso, siendo el patrón hombre la base de la mayoría de ellos. En los últimos 20 años ha existido un vertiginoso desarrollo en el conocimiento acerca del fenómeno de los "cerebros asimétricos". El uso preferencial de uno de un par de órganos simétricos como expresión de una preferencia lateral debida a la organización del cerebro, ha interesado ampliamente a los neurofisiólogos. Existen evidencias abrumadoras de diferencias izquierda-derecha en neuroanatomía y el procesamiento neural de vertebrados e incluso en algunas especies de invertebrados (Halpern, 2005). Numerosos criterios han sido utilizados para caracterizar la dirección de la lateralización. Elementos como la extremidad preferida para golpear, el ojo director, la forma de doblar los brazos: izquierdo sobre derecho o derecho sobre izquierdo y el orden en que se colocan los dedos al cruzar las manos, han sido reconocidos como indicadores de dirección de la lateralización (Mohr et al., 2006; Choudhary y Garg, 2015). El sexo también puede influenciar los patrones de preferencia por el lado derecho o el izquierdo y por ejemplo meta-análisis en humanos adultos han mostrado una mayor prevalencia de zurdos en hombres que en mujeres (Sommer et al., 2008).
Actualmente se reconocen las bases, mediadas por la lateralización, que afectan el comportamiento cotidiano en ambientes silvestres, de una gran cantidad de organismos (Bonati y Csermely, 2013 et al., 2016). La respuesta al ataque de predadores es lateralizada en muchas especies. Los equinos, por ejemplo, responden con más eficiencia cuando el atacante aparece en el campo visual izquierdo (Austin y Rogers, 2014). Esta misma respuesta se evidencia en otros animales (Maciejewska et al., 2016; Schnell et al., 2016), lo que sugiere que el hemisferio derecho del cerebro es el encargado de procesar la información de escape y el miedo condicionado (Rogers et al., 2013). De modo general se ha dicho que el hemisferio derecho, atiende a los estímulos inesperados, asigna propiedades de los objetos y de los alrededores, se distrae fácilmente, observa pequeñas diferencias entre estímulos, responde a las señales globales/geométricas, responde a estímulos, expresa emoción intensa (especialmente de agresión y miedo), detecta y responde para escapar de depredadores, permite el reconocimiento facial y la cognición social. Mientras que el hemisferio izquierdo establece patrones de comportamiento en circunstancias familiares, presta atención enfocada a objetivos o señales específicos, da respuesta sostenida sin distracción, categoriza objetos y otros estímulos, responde a señales de referencia, atiende a procesos guiados por instrucciones aprendidas y al procesamiento secuencial (Rogers et al., 2013).
Un cerebro lateralizado puede conferir ventajas cruciales, lo que permite economizar tejido neural al evitar la duplicación de funciones en los dos hemisferios (Levy, 1977), procesando información en paralelo y evitando el inicio de respuestas simultáneas opuestas. Todo esto al hacer que un hemisferio tenga el control de las acciones (especialmente en animales con órganos sensores situados lateralmente) (Vallortigara, 2000). Adicionalmente, Rogers (2000) sugirió que el incremento de las habilidades cognitivas es uno de los beneficios potenciales de la lateralización cerebral, puesto que animales con cerebros fuertemente lateralizados, pueden tener la habilidad de actuar directamente ante muchas fuentes de información al mismo tiempo. Esto mejora las capacidades para buscar alimento y responder al ambiente.
No obstante, los argumentos acerca de que la lateralización incrementa la eficiencia individual no explican la lateralización a nivel poblacional; la cual puede presentar desventajas ecológicas puesto que hace más predecible el comportamiento individual para sus depredadores (Frasnelli, 2013). Sin embargo, el punto crucial es que todas las ventajas resultantes de la lateralización podrían obtenerse y mantenerse dentro de los cerebros asimétricos individualmente, sin necesidad de alinear la dirección de las asimetrías a escala de la población (Vallortigara, 2000). La conducta de escape lateralizada a nivel poblacional puede ser ventajosa para organismos sociales que escapan al unísono, confundiendo a sus depredadores, pero dado que los comportamientos sociales constituyen etapas avanzadas de la evolución, no se entiende claramente cómo evoluciona este comportamiento en organismos no sociales. Los modelos teóricos de evolución de la lateralización sugieren que el alineamiento de la lateralización a escala de población puede haber evolucionado como una "estrategia evolutiva" para que organismos asimétricos coordinaran su comportamiento (Vallortigara y Rogers, 2005). Esta hipótesis conduce a la predicción de que inicialmente, solo los organismos "sociales" pueden haber comenzado a estar lateralizados, mientras los organismos "solitarios" mantendrían lateralización solo al nivel de individuo (Frasnelli, 2013).
Sorprendentemente, no solo en vertebrados un cerebro asimétrico es crucial para el desarrollo de habilidades cognitivas. Recientemente también se han encontrado cerebros asimétricos en invertebrados. La mosca de la fruta (Drosophila melanogaster Meigen) y el nemátodo Caenorhabditis elegans Maupas, son dos ejemplos de invertebrados en los que la asimetría a escala de individuo es necesaria para tener algunas habilidades cognitivas y puede conferir ventajas (Frasnelli, 2013).
Los invertebrados y particularmente los insectos pueden ser excelentes modelos para probar las hipótesis predichas por los modelos teóricos; que proponen a la evolución de la lateralización y la simetría direccional a escala de población, como características a ser encontradas sólo en especies sociales cooperativas (Anfora et al., 2010). En los insectos se puede distinguir con relativa precisión entre comportamiento gregario o solitario y es poco probable que hayan tenido cambios evolutivos mayores en lo que respecta a la socialización (Frasnelli, 2013).
Como contribución a esta línea de conocimiento, el objetivo principal de este trabajo es comprobar la existencia de lateralización a nivel poblacional en la especie Dysdercus andreae L. (Heteroptera: Pyrrhocoridae). Esta especie forma grandes grupos al alimentarse, principalmente de semillas (Panizzi y Grazia, 2015). Desde el punto de vista teórico resulta interesante comprobar la existencia de lateralización en esta especie, si se tiene en cuenta que forma acumulaciones de varios cientos de individuos, lo que indica cierto grado de coordinación, pero no es estrictamente una especie social.
MATERIALES Y MÉTODOS
El trabajo experimental se realizó en los laboratorios de la Facultad de Ciencias Agropecuarias (FCA) de la Universidad Central "Marta Abreu" de Las Villas (UCLV). La especie utilizada fue Dysdercus andreae L., (Artrhopoda; Insecta; Heteroptera: Pyrrhocoridae). Se seleccionó por presentar una conducta activa, son ágiles, responden rápidamente ante estímulos y no son agresivos ante la manipulación. Además, abundan en cierta época del año, lo que permite obtener cientos de individuos experimentales. Se encuentran formando grupos muy numerosos para alimentarse (principalmente de semillas) y copular, lo que indica cierto grado de coordinación entre individuos (Figura 1).
Se incluyeron 426 individuos adultos, los cuales fueron colectados en áreas del Jardín Botánico de Villa Clara, situado en la UCLV. Para dar respuesta al objetivo trazado se evaluaron cuatro variables a cada individuo:
* Variable 1 - Dirección de respuesta sin estrés
* Variable 2 - Dirección de respuesta de escape
* Variable 3 - Solapamiento de alas
* Variable 4 - Sexo
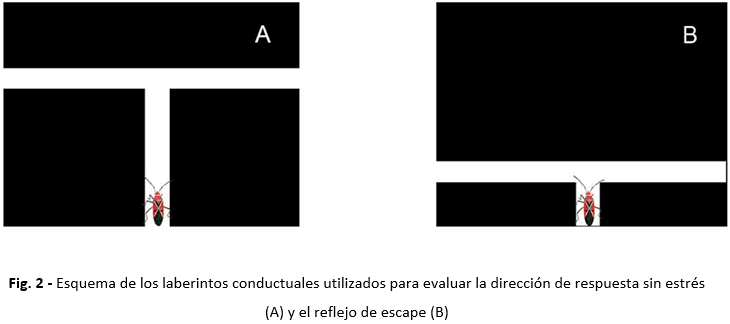
Para la obtención de valores correspondientes a las variables 1 y 2 se diseñaron y construyeron dos laberintos conductuales (Figura 2).
* Laberinto A (para evaluar respuesta sin estrés): Se constituyó con una plataforma de madera de nueve x nueve centímetros, en la cual se trazó un recorrido en forma de letra T, aperturas a tres lados, una sección de entrada y dos de escape opuestas entre sí y perpendiculares a la de entrada. La sección de entrada tenía una longitud de más de tres veces el largo del animal (alrededor de 5 cm). De este modo se dio al insecto tiempo para procesar la información, antes de decidir doblar a la derecha o a la izquierda al final del recorrido de entrada.
* Laberinto B (para evaluar reflejo de escape): Este poseía características constructivas parecidas al laberinto A, excepto que la sección de entrada solo tenía 1,5 cm de longitud, ajustada al largo del cuerpo del insecto, con la intención de que al momento de decidir su dirección de respuesta (derecha o izquierda), el insecto no tuviera un tiempo previo para la integración neural de la información.
Ambos laberintos estaban cubiertos por una sección de plástico traslúcido que permitió visualizar la respuesta del animal a distancia, y estaban pintados de color negro para lograr un patrón regular que no influyese en las decisiones de los insectos.
Procedimiento experimental
Se tomaron individuos al azar, sujetándolos con una pinza blanda por el extremo posterior del abdomen y se colocaron en el extremo anterior de la sección de entrada de los laberintos, con la cabeza apuntando hacia el interior de estos.
En el laberinto A, el animal fue liberado rápidamente dejando de hacer presión con la pinza y se permitió que realizara libremente el recorrido escogido, registrando si doblaba a la izquierda o a la derecha. En el laberinto B, antes de ser liberado, el animal fue estimulado por corrientes de aire suministrados por una pera desde una distancia de 10 – 15 cm, para generar más estrés, y posteriormente se liberó y dejó que realizara libremente el recorrido de su elección.
Como unidades de respuesta se tomaron derecha (D) e izquierda (I) en correspondencia con la dirección tomada. Se consideró D a la derecha del insecto ubicado con la cabeza hacia el interior del laberinto e I a la dirección opuesta.
Luego del recorrido de cada ejemplar los túneles fueron limpiados con un algodón embebido en alcohol al 70 % para evitar la persistencia de marcas olorosas que pudieran influenciar los recorridos de los ejemplares siguientes y se secaron cuidadosamente antes de colocar un nuevo individuo.
Los individuos que presentaron anomalías morfológicas fueron desechados, al igual que las observaciones en donde los ejemplares caminaron con el dorso hacia abajo en el túnel. En los casos en que los animales regresaban luego de seleccionar una dirección se registró como observación la primera decisión.
Se evaluó a cada uno de los individuos sólo una vez en cada túnel; probando primero su comportamiento en el laberinto A, para reducir el estrés previo a esta evaluación, y luego en el B. La posición de los observadores y de los instrumentos de experimentación se cambió periódicamente alrededor de la mesa de experimentación, para evitar la influencia de otros estímulos sensoriales no detectados en la decisión de los animales.
Después de evaluar los individuos en los túneles, cada uno fue sexado y se registró la posición de solapamiento de las alas. Se denominó derechos a los individuos que tenían el ala derecha sobre la izquierda y zurdos a los que presentaban el ala izquierda sobre la derecha. Los resultados de las observaciones fueron procesados estadísticamente utilizando tablas de contingencia y Test Exacto de Fisher. Como indicadores de correlación se calcularon los coeficientes Phi, Lambda simétrico y Lambda considerando una variable dependiente, acompañados del correspondiente valor de significación estadística.
RESULTADOS Y DISCUSIÓN
Los insectos colocados en el laberinto A, para evaluar la dirección conductual sin estrés, caminaron más pausadamente por el túnel y demoraron en ocasiones de uno a tres segundos antes de decidir por una dirección; mientras que en el laberinto B los individuos partían rápidamente en una dirección u otra (Tabla). Estos comportamientos son indicadores de que los laberintos funcionaron adecuadamente para inducir respuestas reflejas al escapar (Laberinto B) o con cierto grado de integración neural (Laberinto A).
Los tamaños de muestra utilizados fueron suficientes para estimar las proporciones con un margen de error inferior al 5 % y un nivel de confianza del 95 %.
Los insectos al responder sin estrés, prefieren doblar hacia la izquierda en cerca de un 60 % de los casos, mostrando evolución hacia una conducta poblacional lateralizada. Mientras que al responder de modo reflejo ante estímulos aversivos escapan indistintamente a la derecha o izquierda.
Estos resultados apuntan a una evolución separada de ambos comportamientos. La evolución distanciada de estos parámetros propicia a los insectos responder coordinadamente y aprovechar las ventajas que confiere un comportamiento lateralizado; y al mismo tiempo evitar la desventaja ecológica que supone una conducta poblacional lateralizada en el momento de escapar individualmente de amenazas. De este modo se combinan en un individuo el incremento de las habilidades cognitivas y la destreza para actuar ante muchas fuentes de información al mismo tiempo y, por ejemplo, buscar alimento, con un comportamiento menos predecible para sus depredadores.
Al analizar la relación entre la dirección de respuesta sin estrés y el solapamiento de las alas (Figura 3), se encontró que los insectos muestran también lateralización a nivel poblacional al plegar las alas, prefiriendo colocar el ala derecha por encima del ala izquierda. No obstante, no fue posible demostrar que la dirección de giro en los túneles estuviese relacionada con la forma de plegar las alas (Coeficiente de correlación Phi < 0,028).
La proporción de sexos encontrada fue de 1:1, sin mostrar relación con ninguna de las otras variables del estudio, por lo que se puede afirmar que las evidencias de lateralización encontradas fueron independientes del sexo de los individuos.
La lateralización encontrada puede considerarse en concordancia al modelo propuesto por Ghirlanda y Vallortigara (2004) en el que el alineamiento de la dirección del comportamiento asimétrico de una especie, a nivel poblacional, puede ser una estrategia evolutiva para que organismos asimétricos puedan coordinarse entre sí. Según estos investigadores la lateralización poblacional se encontraría típicamente entre las especies sociales. Como se ha dicho anteriormente, la especie estudiada no puede considerarse una especie social en el más estricto sentido, pero el grado de lateralización poblacional encontrado podría ser una respuesta evolutiva que permite coordinar acciones entre los individuos puesto que si es una especie que forma grupos.
Por otra parte, se encontró que existe una correlación entre la dirección de respuesta sin estrés y la dirección de escape. Alrededor de un 65 % de los individuos giraron en la misma dirección al reaccionar sin estrés, o sea con integración neuronal, que al reaccionar intuitivamente para escapar (Figura 4). Este resultado es interesante si se tiene en cuenta que; en este estudio la conducta de escape (refleja) no se encontró lateralizada a escala de población, pero si la respuesta con integración neural.
Esta correlación permite enriquecer el modelo evolutivo proponiendo que la lateralización poblacional se desarrolla en un inicio para respuestas más complejas e influye progresivamente sobre la conducta intuitiva de escape; conduciendo también hacia la lateralización poblacional de esta. De este modo se puede explicar el que evolucione una respuesta de escape lateralizada en especies no sociales y que luego se instaure como carácter exitoso en especies que forman grupos. Una conducta de escape lateralizada a nivel poblacional, puede constituir una desventaja para organismos individuales al hacerlos más predecibles para sus depredadores. Sin embargo, puede constituir una ventaja a la hora de evadir depredadores en especies gregarias, como se presenta por ejemplo en algunas especies de peces que forman cardúmenes. En estas condiciones una conducta de escape lateralizada podría convertirse en un comportamiento exitoso y fijarse como elemento evolutivo.
Obviamente, este tipo de hipótesis no puede ser demostrada o refutada en el marco de este único experimento. Para ello sería necesario realizar estudios comparativos de comportamiento con un número representativo de especies. La mayor disponibilidad de datos comparativos sería crucial para el desarrollo de nuestra comprensión de los orígenes evolutivos de la lateralización cerebral.
CONCLUSIONES
1. Existe una conducta poblacional lateralizada en insectos de la especie Dysdercus andreae.
2. La respuesta de escape y las decisiones sin estrés son comportamientos que evolucionaron separadamente en Dysdercus andreae, los individuos prefirieron la izquierda al reaccionar con integración neural, mientras que al responder de modo reflejo a estímulos aversivos escaparon indistintamente a la derecha o izquierda.
3. Se encontró correlación entre la dirección de respuesta sin estrés y la dirección de escape en Dysdercus andreae, lo que puede ser indicativo de que la lateralización poblacional existente en un inicio para respuestas más complejas, influye progresivamente sobre la conducta intuitiva de escape; conduciendo también hacia la lateralización poblacional de esta.
4. Las evidencias de lateralización encontradas fueron independientes del sexo de los individuos.
BIBLIOGRAFÍA
ANFORA, G., FRASNELLI, E., MACCAGNANI, B., ROGERS, L.J. and VALLORTIGARA, G. 2010. Behavioural and electrophysiological lateralization in a social (Apis mellifera) but not in a non-social (Osmia cornuta) species of bee. Behavioural Brain Research, 206 (2): 236-239. DOI 10.1016/j.bbr.2009.09.023. Disponible en: http://www.sciencedirect.com/science/article/pii/S0166432809005385.
AUSTIN, N.P. and ROGERS, L.J. 2014. Lateralization of agonistic and vigilance responses in Przewalski horses (Equus przewalskii). Applied Animal Behaviour Science, vol. 151: 43-50. DOI 10.1016/j.applanim.2013.11.011. Disponible en: http://dx.doi.org/10.1016/j.applanim.2013.11.011.
BONATI, B. and CSERMELY, D. 2013. Lateralization in Lizards: Evidence of Presence in Several Contexts. En: D. CSERMELY y L. REGOLIN (eds.), Behavioral Lateralization in Vertebrates: Two Sides of the Same Coin. Berlin, Heidelberg: Springer Berlin Heidelberg, pp. 25-38. ISBN 978-3-642-30203-9. Disponible en: http://dx.doi.org/10.1007/978-3-642-30203-9_3.
BROCA, P. 1865. Sur le siège de la faculté du langage articulé. Bulletins de la Société d'anthropologie de Paris, 6 (1): 377-393. DOI 10.3406/bmsap.1865.9495. Disponible en: http://www.persee.fr/web/revues/home/prescript/article/bmsap_0301-8644_1865_num_6_1_9495.
CHOUDHARY, A. and GARG, N. 2015. Distribution of Lateralities in Study Population. Indian Journal of Clinical Anatomy and Physiology, 2 (June): 81-83.
FRASNELLI, E. 2013. Brain and behavioral lateralization in invertebrates. Frontiers in Psychology, 4 (DEC): 939. DOI 10.3389/fpsyg.2013.00939. Disponible en: http://journal.frontiersin.org/article/10.3389/fpsyg.2013.00939/abstract.
GHIRLANDA, S. and VALLORTIGARA, G. 2004. The Evolution of Brain Lateralization: A Game-Theoretical Analysis of Population Structure The evolution of brain lateralization: a game- theoretical analysis of population structure. The Royal Society, 271 (1541): 853-857. DOI 10.1098/rspb.2003.2669. Disponible en: http://www.jstor.org/stable/4142518.
HALPERN, M.E. 2005. Lateralization of the Vertebrate Brain: Taking the Side of Model Systems. Journal of Neuroscience, 25 (45): 10351-10357. DOI 10.1523/JNEUROSCI.3439-05.2005. Disponible en: http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.3439-05.2005.
LEVY, J. 1977. The mammalian brain and the adaptive advantage of cerebral asymmetry. Annals of the New York Academy of Sciences, 299 (1): 264-72. DOI 10.1111/j.1749-6632.1977.tb41913.x. Disponible en: http://onlinelibrary.wiley.com/doi/10.1111/j.1749-6632.1977.tb41913.x/pdf.
MACIEJEWSKA, M., ZIÄBA, K., SZYMAÅSKA, J. and WAROÅSKA, M. 2016. Spontaneous laterality in mouse Crl:CD1. Laterality: Asymmetries of Body, Brain and Cognition, 21 (1): 76-84. DOI 10.1080/1357650X.2015.1075024. Disponible en: http://www.tandfonline.com/doi/full/10.1080/1357650X.2015.1075024.
MOHR, C., THUT, G., LANDIS, T. and BRUGGER, P. 2006. Arm folding, hand clasping, and Luria's concept of «latent left-handedness». Laterality: Asymmetries of Body, Brain and Cognition, 11 (1): 15-32. DOI 10.1080/13576500500199795. Disponible en: http://www.tandfonline.com/doi/abs/10.1080/13576500500199795.
PANIZZI, A.R. and GRAZIA, J. (eds.). 2015. True Bugs (Heteroptera) of the Neotropics. Dordrecht: Springer Netherlands. ISBN 978-94-017-9860-0. Disponible en: http://link.springer.com/10.1007/978-94-017-9861-7_6\nhttp://link.springer.com/10.1007/978-94-017-9861-7.
PAOLI, M., ANDRIONE, M. and HAASE, A. 2016. Imaging techniques for the study of lateralization in insects. En: L.J. ROGERS y G. VALLORTIGARA (eds.), Lateralized brain functions. New York: Springer New York, pp. 695. ISBN 978-1-4939-6725-4. Disponible en: http://libgen.io/get/585712A79EE3D37E9CF87DA43F255AAA/.
ROGERS, L.J. 2000. Evolution of Hemispheric Specialization: Advantages and Disadvantages. Brain and Language, 73 (2): 236-253. DOI 10.1006/brln.2000.2305. Disponible en: http://www.sciencedirect.com/science/article/pii/S0093934X00923058.
ROGERS, L.J., VALLORTIGARA, G. and ANDREW, R.J. 2013. Divided brains. The biology and behaviour of brain asymmetries. New York: Cambridge University Press. ISBN 978-0-521-18304-8.
SCHNELL, A.K., HANLON, R.T., BENKADA, A. and JOZET-ALVES, C. 2016. Lateralization of Eye Use in Cuttlefish: Opposite Direction for Anti-Predatory and Predatory Behaviors. Frontiers in Physiology, 7 (December): 1-8. DOI 10.3389/fphys.2016.00620. Disponible en: http://journal.frontiersin.org/article/10.3389/fphys.2016.00620/full.
SOMMER, I.E., ALEMAN, A., SOMERS, M., BOKS, M.P. and KAHN, R.S. 2008. Sex differences in handedness, asymmetry of the Planum Temporale and functional language lateralization. Brain Research, 1206 (C): 76-88. DOI 10.1016/j.brainres.2008.01.003. Disponible en: http://linkinghub.elsevier.com/retrieve/pii/S0006899308000401.
VALLORTIGARA, G. 2000. Comparative Neuropsychology of the Dual Brain: A Stroll through Animals' Left and Right Perceptual Worlds. Brain and Language, 73 (2): 189-219. DOI 10.1006/brln.2000.2303. Disponible en: http://www.ibrarian.net/navon/paper/Comparative_Neuropsychology_of_the_Dual_Brain__A_.pdf?paperid=2327052.
VALLORTIGARA, G. and ROGERS, L.J. 2005. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behavioral and Brain Science, 28 (4): 575-89. DOI 10.1017/S0140525X05000105.
Recibido el el 11 de mayo de 2017 y aceptado el 19 de septiembre de 2018
{kind=link}