ARTICULOS GENERALES
Efecto de la salinidad sobre el contenido relativo de agua y la concentración de pigmentos en tres genotipos de frijol (Phaseolus vulgaris L.)
Effect of salinity on relative water content and pigment concentration in three genotypes of bean (Phaseolus vulgaris L.)
Licet Chávez Suárez, Alexander Álvarez Fonseca, Ramiro Ramírez Fernández, Sucel Infante Fonseca, Luis Licea Castro, Blanca García Rodríguez, Aida García Alcántara y Milvia Fonseca Arias
Instituto de Investigaciones Agropecuarias
“Jorge Dimitrov”. Carretera Vía Manzanillo Km 17. Bayamo. Granma.
Cuba, C.P. 85100.
E-mail: licet@dimitrov.cu
RESUMEN
El presente trabajo se desarrolló en el Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, en el mes de noviembre de 2011, con el objetivo de estudiar el efecto de la salinidad sobre el contenido relativo de agua (CRA) y la concentración de pigmentos, en plantas de tres variedades de frijol (Phaseolus vulgaris L.): HOL-17, HOL-35, HOL-74, utilizando un suelo salino (CE 5,2 dS.m-1) y un suelo no salino como control. Se determinó el contenido relativo de agua (CRA) a través del método descrito por Yamasaki y Rebello y la concentración de pigmentos según la metodología de Lichtenthaler y Wellburn. Los resultados indicaron que disminuyó significativamente el CRA en el tratamiento salino respecto al control, en las tres variedades evaluadas. El contenido de clorofila b se afectó en las tres variedades cuando se sometieron a las condiciones salinas. Las variedades HOL-74 y HOL-17 exhibieron un incremento significativo en la concentración de carotenoides, lo que sugiere un mecanismo para proteger la clorofila de la fotooxidación provocada por el estrés salino.
Palabras clave: frijol, estrés salino, pigmentos, contenido relativo de agua.
ABSTRACT
This work was developed at the Agricultural Research Institute “Jorge Dimitrov” in November 2011, with the aim of studying the effect of salinity on the relative water content (RWC) and the concentration of pigments in plants of three varieties of beans (Phaseolus vulgaris L.): HOL-17 HOL-35 HOL-74, using a saline soil (EC 5.2 dS.m-1) and a non-saline soil as control. The Relative Water Content (RWC) was determined by the method described by Yamasaki and Rebello and the concentration of pigment by the methodology of Lichtenthaler and Wellburn. The results indicated a significant decrease in the RWC in the saline treatment, relative to control, in the three studied varieties. The content of chlorophyll b was affected in the three varieties under saline conditions, while HOL-17 and HOL-74 varieties exhibited a significant increase in the concentration of carotenoids, suggesting a protective mechanism of chlorophyll photooxidation induced by salt stress.
Key words: bean, saline stress, pigment, relative water content.
INTRODUCCIÓN
La salinidad es un estrés abiótico complejo que simultáneamente presenta componentes osmóticos e iónicos. La pérdida de la homeostasis, tanto hídrica como iónica, ocurre tanto a escala celular como a nivel de planta y provoca graves daños moleculares que detienen el crecimiento. Por una parte, la disminución del potencial hídrico del medio restringe la absorción de agua por las raíces y por otra, los iones salinos se acumulan dentro de los tejidos de la planta en concentraciones que llegan a ser tóxicas y pueden, al mismo tiempo, inducir desequilibrios nutricionales por modificación de la absorción y distribución de nutrientes esenciales (Morales et al., 2010).
El hecho de que en condiciones salinas, la disponibilidad de agua para las plantas disminuye y estas se vean obligadas a vivir en condiciones de sequía fisiológica, ha motivado el desarrollo de un número elevado de investigaciones encaminadas a dilucidar el efecto de la salinidad en sus relaciones hídricas (González, 2001). Por otro lado, la salinidad afecta la fotosíntesis, principalmente a través de la reducción del área foliar, el contenido de clorofila, la conductancia estomática, y en menor extensión, a través de una disminución de la eficiencia del fotosistema II (Chaves, 2009).
Es objetivo del
presente trabajo es evaluar el efecto de la salinidad sobre el contenido relativo
de agua y la concentración de pigmentos en plantas de tres variedades
de frijol común (Phaseolus vulgaris L.).
MATERIALES Y MÉTODOS
El experimento se realizó en el laboratorio y la casa con techo de cristal del Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, municipio de Bayamo, Granma, en el mes de noviembre del 2011.
Se seleccionaron semillas de tres variedades de frijol común (HOL-17, HOL-35 y HOL-74) provenientes del banco de germoplasma del Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, con más de un 90 % de germinación.
Se sembraron dos semillas de estas variedades en bolsas de polietileno de 1 Kg de capacidad. Para cada variedad se establecieron dos variantes experimentales: un tratamiento salino que consistió en la utilización de un suelo salino con una conductividad eléctrica medida por pasta de saturación de 5,2 dS.m-1 y como control, un suelo no salino que presentó características similares. Se utilizó un diseño completamente aleatorizado y por cada tratamiento se utilizaron cuatro réplicas de 20 bolsas cada una.
27 días después de la siembra se muestrearon 10 plantas por réplica a las 7 a.m., para un total de 40 plantas por tratamiento a las que se le determinaron el CRA de acuerdo con el procedimiento de medición por gravimetría descrito por Yamasaki y Rebello (1999), utilizando la siguiente ecuación:
CRA (%) = [(MF – MS)/(MT- MS)]x100
Donde:
· MF- Masa fresca
· MS- Masa seca
· MT- Masa turgente
Para determinar la concentración de pigmentos (clorofila a, b y carotenos)
se siguió la metodología de Lichtenthaler y Wellburn (1984). Se
realizaron cinco repeticiones. Para ello se pesó un gramo de tejido foliar
y después de cortado en pequeños pedazos, se maceraron con alcohol
etílico (96 v/v), filtraron a través de un papel de filtro de
poros finos, y completado el contenido hasta un volumen de 50 mL con alcohol
etílico. Posteriormente fue leída la absorbancia a 665, 649 y
470 nm para la clorofila a (Cla), b (Clb) y los carotenos respectivamente, e
inmediatamente se calculó la concentración de pigmentos en µg
por cada ml de extracto-1, según las ecuaciones siguientes:
Cla = 13.95 A665
– 6.68 A649
Clb = 24.96 A649 – 7.32 A665
Cx+c = (1000 A470 – 2.05 Cla – 114.8 Clb) / 245
Donde:
A470 , A649, A665 – absorbancia a 470, 649 y 665 nm
Adicionalmente se determinó la relación Cla/Clb.
Al determinar la media y el error estándar para los indicadores evaluados
en ambas condiciones se establecieron las diferencias entre el tratamiento control
y de estrés para cada variedad, mediante la prueba t de Student (p<0,05),
utilizando paquete estadístico “Statistics ver. 8 para Windows.
Previamente, se comprobó la normalidad y la homogeneidad de varianzas
de los datos mediante las pruebas de Kolmogorov– Smirnov y de Bartlet (Cochran
y Cox, 1992).
RESULTADOS Y DISCUSIÓN
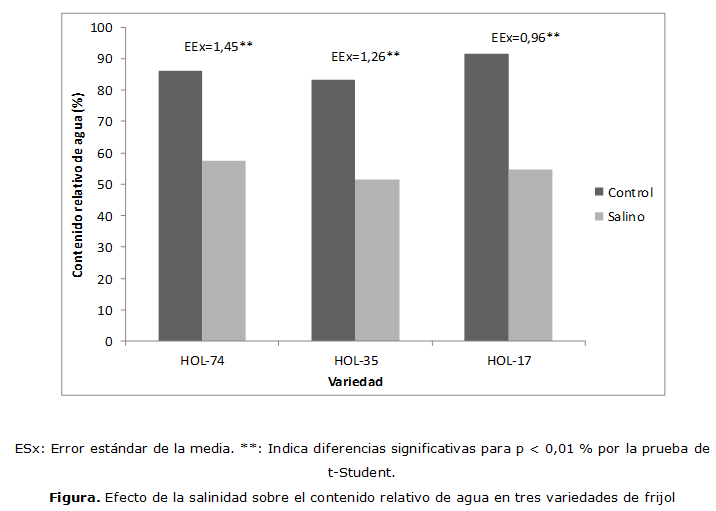
Como se muestra en la (Figura), el CRA en las variedades estudiadas disminuyó de forma significativa en el tratamiento salino respecto al control. Este resultado demuestra la ineficiente economía del agua en las plantas cultivadas bajo condiciones salinas debido a la aparición de un estado de sequía fisiológica. Este fenómeno no se debe a la falta de agua en el suelo, sino a que la planta no puede absorberla con facilidad porque el suelo posee una alta concentración de sales y por tanto valores del potencial hídrico muy bajos que pueden llegar a ser inferiores al potencial de la célula, limitando la absorción del agua (González, 2001).
Hossain et al. (2011) señalaron una disminución significativa en el CRA, con el incremento de la concentración salina, en la especie Arachis hipogea L. Esta disminución fue del 13 % comparado con el control a la concentración de 200 mM de NaCl. Lefi y Ben (2014), por su parte, observaron una disminución en este indicador al evaluar el comportamiento en condiciones salinas de dos especies del género Pistacia.
En genotipos de trigo, Abdelmalek y Khaled (2011) constataron la reducción significativa del CRA (24 %), con los niveles de salinidad aplicados y la correlacionaron con la disminución del potencial hídrico y el potencial osmótico, verificados en el experimento. Asimismo, Akbari et al. (2012) observaron una disminución significativa del CRA al aplicar concentraciones salinas 5, 10, 15 dS m-1 en condiciones de invernadero a cuatro variedades de trigo.
De igual forma Shanker et al. (2014) constataron una reducción significativa de este indicador en concentraciones salinas mayores que 3 dS m-1 al evaluar el comportamiento de cinco cultivares de (Brassica juncea L.); mientras Quin et al. (2010) en plántulas de Shepherdia argéntea (Pursh) Nutt., observaron una disminución para las concentraciones salinas de 400 y 600 mmol/L.
Sin embargo, no existen diferencias significativas entre las concentraciones de clorofila a en el tratamiento salino respecto al control, en las variedades HOL-74 y HOL-35.
Al evaluar la clorofila b se observó una reducción significativa de la misma en las plantas cultivadas bajo estrés salino y un incremento de la relación Cla/ Clb para las tres variedades. No obstante, es preciso apuntar que los valores de la relación Cla/Clb coinciden con el rango señalado (1- 3) por Srivastava (1998) para las plantas C3.
Además, se registró un incremento significativo en la concentración de carotenoides de las plantas cultivadas en condiciones salinas respecto al tratamiento control en las variedades HOL-74 y HOL-17, mientras que la variedad HOL-35 no mostró diferencias con el tratamiento control (Tabla).
El contenido de clorofila desempeña un papel importante en el crecimiento y desarrollo de las plantas (Jahan et al., 2014) y la disminución en su contenido es la principal causa de alteraciones en el proceso de fotosíntesis (Khairi et al., 2015). En este sentido, Torres et al. (2005) refieren que una de las causas de la disminución de la intensidad de la fotosíntesis bajo estrés salino, es la disminución de la concentración de pigmentos en las hojas.
García et al. (2010), al evaluar dos cultivares de Phaseolus, encontraron una disminución del contenido relativo de clorofila y atribuyeron esta reducción a la inhibición en la síntesis de precursores de esta molécula lo que, por lo común, se acentúa a medida que se prolonga el período de estrés.
Akca y Samsunlu (2012) constataron una disminución significativa en el contenido de clorofilas a y b en plantas de nogal (Jungla regia L.), sometidas a tratamiento salino, lo que estuvo en dependencia de los niveles de salinidad aplicados con el riego. A la vez, Ghassemi y Mahmoodi (2012) obtuvieron resultados similares al evaluar el comportamiento de plantas de lenteja (Lens culinaris Medik.).
Hossain et al. (2011), en A. hipogea, constataron una disminución del 50 % del contendido de clorofila total con la utilización de concentraciones salinas de 200 mM de NaCl, y lo atribuyeron a daños de los cloroplastos provocados por el estrés salino. Por su parte, Gomathi y Rakkiyapan (2011) al evaluar el efecto del estrés salino en genotipos de caña de azúcar, observaron una disminución significativa del contenido de clorofila y carotenoides en diferentes estadios de desarrollo del cultivo.
Stepien y Johnson (2009) señalaron que la concentración de clorofila se ve afectada significativamente en Arabidopsis thaliana (L.) Heynh. pero no en Thellungiella sp., especie que se ha utilizado como modelo de cultivo tolerante a la salinidad.
En contraste Higbie et al. (2010) refieren que genotipos de Gossypium sp. incrementaron significativamente el contenido de clorofila cuando se encontraron en condiciones salinas, mientras que no se afectó la emisión de fluorescencia. No obstante, Akbari et al. (2012), observaron un incremento de la concentración de clorofila durante la fase vegetativa en cuatro cultivares de trigo y la disminución de este indicador durante la fase de floración, bajo condiciones de salinidad.
Por otro lado, el incremento del contenido de carotenoides pudiera sugerir un mecanismo de adaptación a estas condiciones, de las variedades HOL-74 y HOL-17, si se tiene en cuenta el papel de los carotenoides en la protección de la molécula de clorofila durante el proceso de fotooxidación. De acuerdo con Zhang y Kirkhen (1996) los carotenoides pueden reaccionar con los estados excitados de la clorofila y combinados con el oxígeno molecular, constituir un complejo que conduce a la fotooxidación.
Una fracción de los carotenoides, la violaxantina, participa en el ciclo de las xantofilas, que se ha propuesto como un mecanismo de protección para las plantas, sobre todo del aparato fotosintético y el daño por radicales libres. De hecho, se ha correlacionado la mayor actividad de este ciclo con la resistencia de las plantas al daño por fotooxidación que se produce bajo el estrés salino o por la sequía (Raya-Pérez, 1998). En este sentido, se puede plantear que la variedad HOL-35, al no exhibir este mecanismo de adaptación, es más susceptible que las otras dos variedades. Sin embargo se sugiere continuar las investigaciones en otras fases del cultivo sobre la base de un mayor número de indicadores fisiológicos y agronómicos.
CONCLUSIONES
1. El CRA disminuyó de forma significativa en el tratamiento salino con relación al control en las tres variedades de frijol evaluadas.
2. El contenido de clorofila b se afectó de forma significativa en las variedades evaluadas.
3. El contenido de carotenoides en las variedades HOL-17 y HOL-74 tuvo un incremento significativo.
BIBLIOGRAFÍA
1. Akbari, E.; A. Izadi; A. Borzouei: Effects of salinity on some physiological traits in wheat (Triticum aestivum L.) cultivars. Indian Journal of Science and Technology 5(1): 1901-1906, 2012.
2. Akca, Y. y E. Samsunlu: The effect of salt stress on growth, chlorophyll content, proline and nutrient accumulation, and K/Na ratio in walnut. Pak. J. Bot., 44 (5): 1513-1520, 2012.
3. Abdelmalek, C. y T. Khaled: Physiological behavior of wheat genotypes from Algerian semi-arid regions grown under salt stress. African Journal of Agricultural Research 5 (23): 636-641, 2011.
4. Chaves, M.; J. Flexas; C. Pinheiro: Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany 103 (4): 551-560, 2009.
5. Cochran, W. y G. Cox: Experimental design. New York. USA. 1992, 640 p. ISBN: 978-0-471-54567-5
6. García, M.; G. García; M. Sanabria: Efecto de la salinidad sobre el crecimiento, daño oxidativo y concentración foliar de metabolitos secundarios en dos variedades de caraota (Phaseolus vulgaris L.). Interciencia 35(11): 840-846, 2010.
7. Ghassemi, K. y F. Mahmoodi: Physiological responses of lentil (Lens culinaris Medik.) to salinity. International Journal of Agriculture and Crop Science 4 (20): 1531-1535, 2012.
8. Gomathi, R. y P. Rakkiyapan: Comparative lipid peroxidation, leaf membrane thermostability, and antioxidant system in four sugarcane genotypes differing in salt tolerance. International Journal of Plant Physiology and Biochemistry 3 (4): 67-74, 2011.
9. González, L.M.: Apuntes sobre la fisiología de las plantas cultivadas en condiciones salinas. Cultivos Tropicales 23 (4):47-57, 2001.
10. Higbie, S.M.; F. Wang; J. Stewart; T. Sterling; W. Lindemann; E. Hughs; J. Zhang: Physiological response to salt (NaCl) stress in selected cultivated tetraploid cottons. International Journal of Agronomy, volumen 10, Article ID 643475, 12 p. en sitio web: http://www.hindawi.com/journals/ija/2010/643475/ consultado el 03 de marzo de 2015. doi:10.1155/2010/643475, 2010.
11. Hossain, M.A.; M. Ashrafuzzaman; M. Ismail: Salinity triggers proline synthesis in peanut leaves. Maejo Int. J. Sci. Technol. 5(01): 159-168, 2011.
12. Jahan, M.S.; M. Nozulaidi; M. Moneruzzaman; A. Ainun; N. Husna: Control of plant growth and water loss by a lack of light-harvesting complexes in photosystem-II in Arabidopsis thaliana ch1-1 mutant. Acta Physiologiae Plantarum 36:1627-1635, 2014.
13. Khairi, M.; M. Nozulaidi; M. Sarwar: Effects of different water levels on physiology and yield of salinity rice variety. Australian Journal of Basic and Applied Sciences 9(2): 339-345, 2015.
14. Lefi, E. y H. Ben: Effects of salt stress on plant water status, leaf gas exchanges and chlorophyll fluorescence of Pistacia atlantica Desf. versus Pistacia vera L. International Journal of Agronomy and Agricultural Research 5(6): 64-77, 2014.
15. Lichtenthaler, H.K. y A.R. Wellburn: Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions 603: 591-592, 1984.
16. Morales, D.; P. Rodríguez; J.M. Dell’Amico; A. Torrecillas; M.J. Sánchez: Efecto del estrés por NaCl en el crecimiento y las relaciones hídricas en plantas de tomate (Solanum lycopersicum L.) durante el período vegetativo. Cultivos Tropicales 31(4): 76-81, 2010.
17. Quin, J.; W.Y. Dong; K.N. He; Y. Yu; G.D. Tan; L. Han; M. Dong; Y. Zhang; D. Zhang; A.Z. Li; Z.L. Wang: NaCl salinity-induced changes in water status, ion contents and photosynthetic properties of Shepherdia argentea (Pursh) Nutt. Seedlings. Plant Soil Environ. 56(7): 325-332, 2010.
18. Raya-Pérez, J.C.: Resistencia a sequía, un avistamiento. Revista Chapingo Serie Ciencias Forestales y del Ambiente 4(2): 267-272, 1998.
19. Shanker, K.; S. Parihar; S. Biswas: Effect of saline water on relative water content, rate of photosynthesis, chlorophyll content and yield of indian mustard (Brassica juncea L.). International Journal of Bio-resource and Stress Management 5(1): 37-40, 2014.
20. Srivastava, M.: Effects if ionizing radiation on Xanthine oxireductase system. Doctor Thesis. Jawaharlal Neru University. New Delhi, 1998, 100 pp.
21. Stepien, P. y G. Johnson. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte Thellungiella: Role of the plastid terminal oxidase as an alternative electron Sink. Plant Physiology 149:1154-1165, 2009.
22. Torres, A.; E. Campostrini; J. Goncalves; R. Bressan-Smith: Photosynthetic pigments, nitrogen, chlorophylla ûuorescence and SPAD-502 readings in coffee leaves. Scientia Horticulturae 104:199-209, 2005.
23. Yamasaki, S. y L. Rebello: Measurement of leaf relative water content in Araucaria angustifolia. Revista Brasileira de Fisiología Vegetal 11(2): 69-75, 1999.
24. Zhang, J. y
M. Kirkhan: Antioxidant responses to drought in sunflower and sorghum seedlings.
New Phytology 132: 361-373, 1996.
Recibido:
06/04/2014
Aceptado: 05/06/2015
{kind=link}
{kind=link}